/ Beyond Nootropics: Why Brain-training Is The Missing Link
Last edited: 2026-01-13
Modern cognitive optimization often centers around improving nutrition, balancing neurotransmitters, or using nootropics to enhance alertness and focus. While these interventions can be useful, their impact is fundamentally limited by a core principle of neuroscience: most cognitive functions follow an inverted U-shaped curve.
Performance is impaired when neuromodulatory activity is too low, impaired again when it is too high, and optimal only within a relatively narrow middle range [1][2]. This observation is ubiquitous in that several distinct neurotransmitter systems, including dopamine, noradrenaline, and acetylcholine, all exhibit this common characteristic. Importantly, the brain is not passive in this process: it is already continuously regulating neuromodulatory tone through homeostatic and feedback mechanisms to keep itself near functional optima [3].
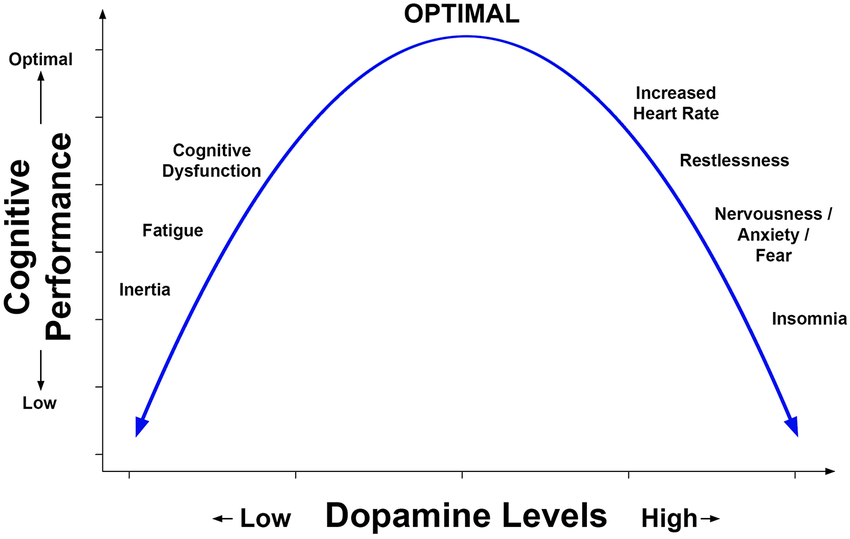
Nootropics and nutritional interventions primarily move you horizontally along the curve. By increasing or decreasing neurotransmitter availability, receptor stimulation, or arousal, they shift your current state left or right. If you start below the optimum—fatigued, under-aroused, or deficient—this can produce noticeable benefits. If you are already near the peak, the same intervention may do little or even impair performance by pushing you past it [4][5]. Crucially, this horizontal shift does not alter the shape of the curve itself. The maximum achievable performance remains fixed, defined by the existing efficiency and capacity of the underlying neural circuits.
Understanding what optimizing this curve can and cannot do makes clear why brain training is the indispensable missing link. Brain training operates on a different axis. Repeated, demanding cognitive tasks drive activity-dependent plasticity in the very networks that implement working memory, attention, and cognitive control. Over time, this can vertically raise the peak height of the curve by increasing representational fidelity, improving signal-to-noise ratios, and strengthening recurrent network dynamics in regions such as the prefrontal cortex and striatum [6–8]. In practical terms, the brain becomes capable of better performance even when neuromodulatory conditions are unchanged.
There is also a plausible, but less firmly established, case that training may broaden the curve, making performance less sensitive to precise neuromodulatory tuning. Stronger, more efficient circuits may require less finely calibrated dopamine or noradrenaline levels to remain stable, increasing robustness to stress, fatigue, or distraction. While indirect evidence supports this idea—such as reduced vulnerability of trained networks to interference or stress-induced collapse—direct demonstrations of curve “widening” are limited, and this claim should be treated as provisional rather than settled [7–9]. What is well supported, however, is that training reduces reliance on moment-to-moment neurochemical optimization by embedding the skill more deeply in network structure.
This distinction explains a common experiential pattern. Nootropics often feel like they improve state: focus feels sharper, effort feels lower, thinking feels smoother. Yet these effects are transient and context-dependent. Brain training, by contrast, often feels effortful and slow to pay off, but produces changes that persist even when arousal or motivation fluctuates. The former improves where you sit on the curve today; the latter changes how high the curve goes tomorrow.
Importantly, this does not make nootropics irrelevant. Horizontal shifts still matter because plasticity itself is gated by neuromodulatory state. Dopamine and noradrenaline strongly influence whether synaptic changes occur at all [2][7]. In this sense, nutrition and nootropics can be seen as enablers of learning, helping place the brain in a zone where training-induced plasticity is possible. But without the structured demands of training, there is no signal telling the brain what to optimize.
In summary, optimizing brain chemistry moves you along a fixed performance landscape. Brain training reshapes that landscape by raising its ceiling and possibly broadening the zone of high performance. While possibly synergistic, brain training is likely indispensable as a priming agent for inducing changes in what the brain is capable of, rather than merely optimizing conditions for performance.
There is an important caveat to the principle described above: the term “nootropic” has come to include nutrients and compounds with structural effects on the brain, such as those supporting myelination or cell membrane synthesis. For rigor, the present discussion should be interpreted as applying primarily to classical nootropic compounds with stimulating or neuromodulatory properties. It is plausible that some compounds exert cognitive effects through mechanisms largely orthogonal to the inverted U-shaped framework.
NOTE: We do NOT endorse any particular nootropic or compound, nor do we think that these compounds are essential to see progress with Bird Back. Always consult your healthcare professional.
– PF
References
[1] Yerkes, R. M., & Dodson, J. D. (1908). The relation of strength of stimulus to rapidity of habit-formation. Journal of Comparative Neurology and Psychology.
[2] Arnsten, A. F. T. (1998). Catecholamine modulation of prefrontal cortical cognitive function. Trends in Cognitive Sciences.
[3] Turrigiano, G. G. (2012). Homeostatic synaptic plasticity: Local and global mechanisms for stabilizing neuronal function. Cold Spring Harbor Perspectives in Biology.
[4] Cools, R., & D’Esposito, M. (2011). Inverted-U–shaped dopamine actions on human working memory and cognitive control. Biological Psychiatry.
[5] Robbins, T. W., & Arnsten, A. F. T. (2009). The neuropsychopharmacology of fronto-executive function. Annual Review of Neuroscience.
[6] Klingberg, T. (2010). Training and plasticity of working memory. Trends in Cognitive Sciences.
[7] Goldman-Rakic, P. S., Muly, E. C., & Williams, G. V. (2000). D1 receptors in prefrontal cells and circuits. Brain Research Reviews.
[8] Dahlin, E., Neely, A. S., Larsson, A., Bäckman, L., & Nyberg, L. (2008). Transfer of learning after updating training mediated by the striatum. Science.
[9] McEwen, B. S., & Morrison, J. H. (2013). The brain on stress: vulnerability and plasticity of the prefrontal cortex. Nature Reviews Neuroscience.